Chemische Evolution

„Woher kommen wir? Wer sind wir? Wohin gehen wir?“ Diese Frage stellte sich nicht nur der französische Maler und Grafiker Eugène Henri Paul Gauguin, während er 1897 in seiner selbst errichteten Hütte auf Tahiti über vier Wochen hinweg das gleichnamige Gemälde schuf. Die Frage danach, wo wir herkommen, ist vermutlich so alt wie die Menschheit selbst.
Im Laufe der Zeit haben sich viele bedeutende Denker über diese Frage den Kopf zerbrochen und haben alle ihre eigenen Erklärungen gefunden, manche mehr, viele davon weniger plausibel. Viele dieser Erklärungen vom damals vorherrschenden religiösen Denken geprägt.
Das Miller-Urey Experiment
{kind=link}
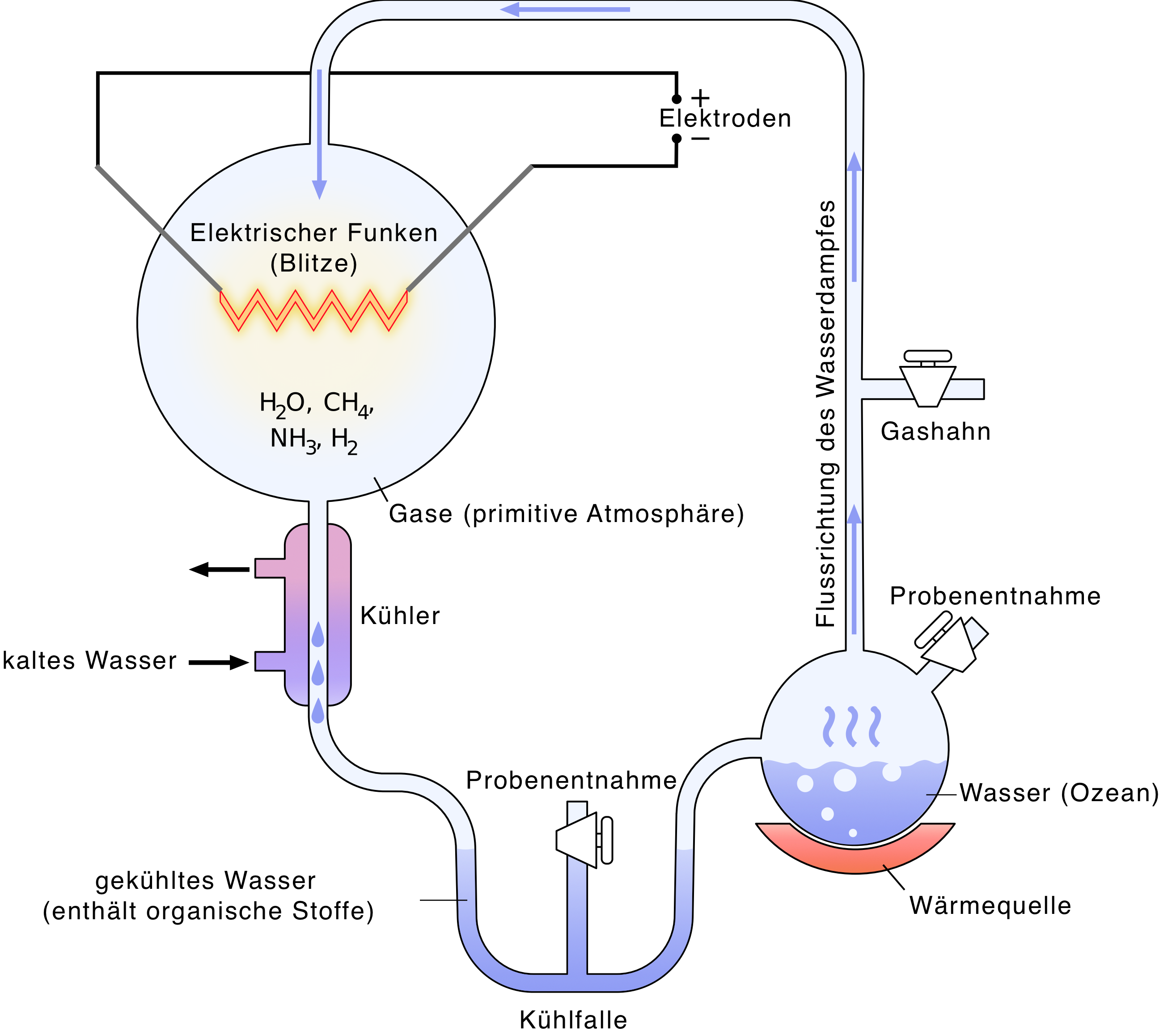
Zusammen mit Harold Clayton Urey führte Stanlay Loyd Miller 1953 ein Experiment durch, welches die Bedingungen auf einer jungen Erde nachstellen sollte. Ziel war es, das Modell der Ursuppe zu untermauern.
Im Versuch wird ein Kolben mit Gasen, welche auf der frühen Erde vermutet werden, gefüllt und diesen durch simulierte Blitze zum Reagieren gebracht. Die entstehenden Produkte werden durch Wasserdampf aufgefangen, welcher kondensiert und aufgefangen wird. Das aufgefangene Wasser wird mit den Reaktionsprodukten wieder in den Verdampfungskolben zurückgeleitet.
Mit diesem sehr einfachen Versuchsaufbau gelang es Miller und Urey, diverse Säuren und Amide herzustellen, darunter auch mehrere Aminosäuren. Moderne Analysen der Rückstände in den Kolben, welche in dem ursprünglichen Experiment verwendet wurden, konnten Spuren von einer weiteren Aminosäure, Ribose, und Adenin nachweisen.
Obwohl die Ergebnisse auch heute noch beeindruckend sind, dafür, dass mit einem so einfachen Versuchsaufbau bereits erste Bausteine von DNA erzeugt werden konnten, äußern sich viele Wissenschaftler dem Versuch kritisch gegenüber. So werden heutzutage Bedingungen auf der jungen Erde vermutet, welche nicht mit denen des Experiments vereinbar sind.
Die Eisen-Schwefel-Welt
Ein moderneres Modell zur Entstehung des Lebens ist die Eisen-Schwefel-Welt. Diese postuliert, dass Leben am Grund der Meere der jungen Erde in den Exhalationen und Aussuppungen von hydrothermalen Schlotten, auch Raucher genannt, entstanden ist. Die hydrothermalen Lösungen, die aus den Schlotten austreten, setzen sich aus gelösten Mineralien und vulkanischen Gasen zusammen und können teils Temperaturen von bis zu 374,15 °C, der kritischen Temperatur von Wasser, erreichen und liefern somit gleichzeitig Material und Energie in Form von Wärme.
Der Name Eisen-Schwefel-Welt leitet sich davon ab, dass Eisen-Schwefel-Mineralien, welche sich um die hydrothermalen Schlotten ablagerten, als Katalysator für die ersten präbiotischen Reaktionen gedient haben sollen und auch sonst im Modell eine zentrale Rolle spielen.
Eisen-Schwefel-Schwämme
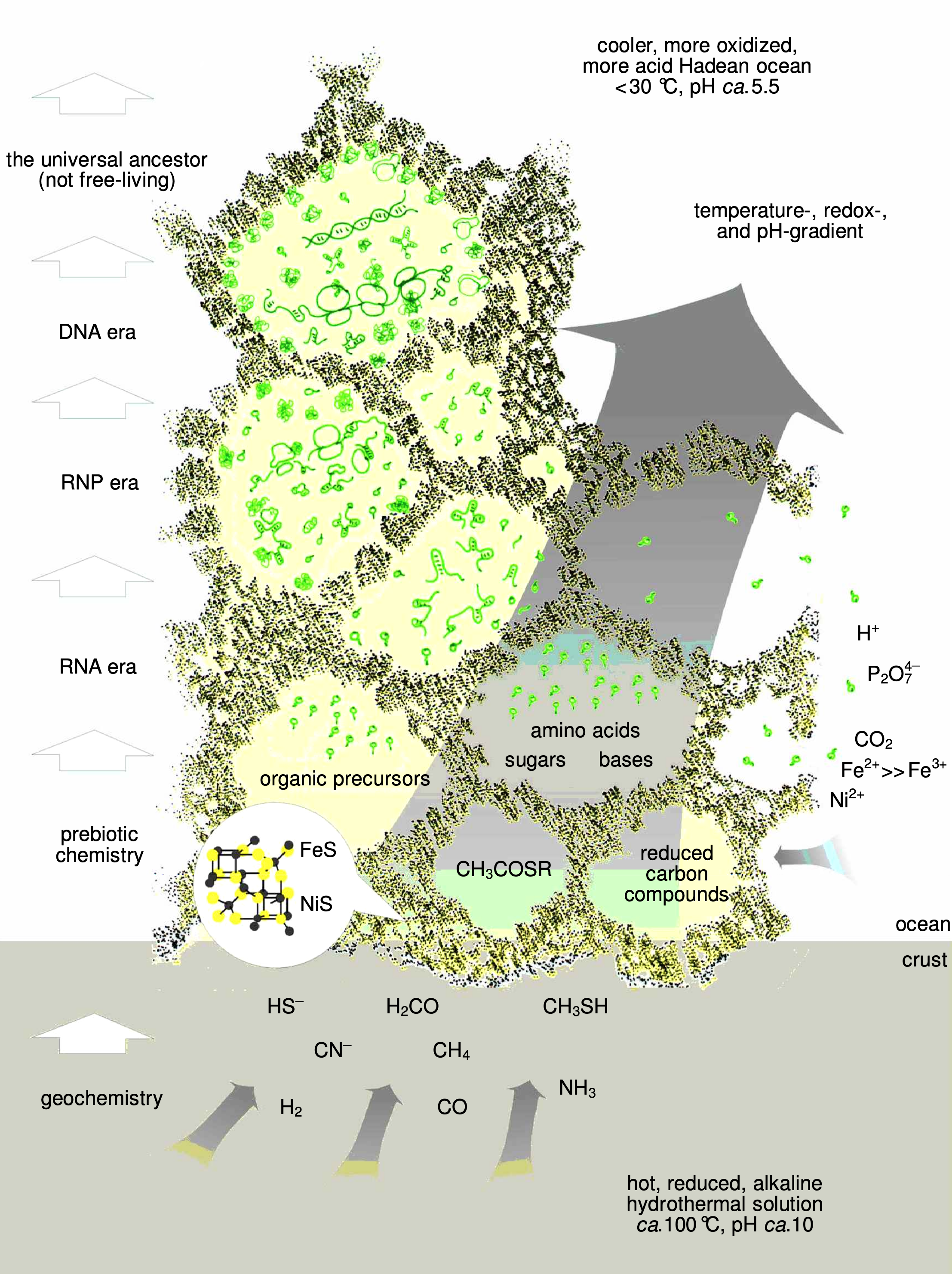
Analysen der Ablagerungen um moderne hydrothermale Schlotten haben ergeben, dass diese nicht ein solider Block, sondern schwammförmig sind. Hauchdünne Wände trennen tausende kleine Poren voneinander. Angenommen, diese Ablagerungen haben sich auch vor 4 Milliarden Jahren formen können, lässt sich ein Modell vorstellen, in welchem die Eisen-Schwefel-Mineralien nicht nur als flache Oberfläche präsent waren, sondern als 3-dimensionale Konstrukte, in deren Poren sich die Moleküle, aus welchen sich später Leben formen würde, sammeln und miteinander reagieren können, wobei diese Reaktionen immer noch von den Eisen-Schwefel-Oberflächen katalysiert werden.
Bereits hier lassen sich Mechanismen der darwinischen Evolution anwenden. Die präbiotisch synthetisierten Nucleobasen, Ribose und Phosphatreste reagieren zu Nucleotiden, welche dann, katalysiert durch die Eisen-Schwefel-Oberflächen, zu Ribozymen (RNA-Moleküle mit katalytischer Aktivität) polymerisieren. Diese agieren wiederum selbst als Katalysator, einerseits für die Synthese von anderen Bio(-makro-)molekülen und für ihre eigene Synthese, womit sie einerseits eine rudimentäre Form eines Stoffwechsels besitzen und sich ebenfalls selbst vermehren können. Die Umweltbedingungen in den Eisen-Schwefel-"Zellen" üben dann einen Evolutionsdruck auf diese aus. Somit haben wir bereits in dieser sehr primitiven Welt von Ribozymen eine Form darwinischer Evolution.
Diese Ribozyme organisieren sich irgendwann zu einem kohärenten "Organismus" innerhalb der Eisen-Schwefel-"Zellmembran". Indem Aminosäuren in die Struktur von Ribonucleinsäuren(RNS) eingebaut werden, formen sich Ribonucleoproteine(RNP), die zusätzliche Möglichkeiten für komplexeren Stoffwechselwege besitzen. Schließlich entwickeln sich im Anschluss an Ribonucleinsäuren(RNS) noch Desoxyribonucleinsäuren(DNS). Diese drei Nucleinsäuren bilden nun das, was man als Vorläufer einer Zelle betrachten kann; jedoch besitzen diese keine Phospholipidmembranen, sondern sind fest in Eisen-Schwefel-"Membranen" beheimatet.
Freilebende Zellen
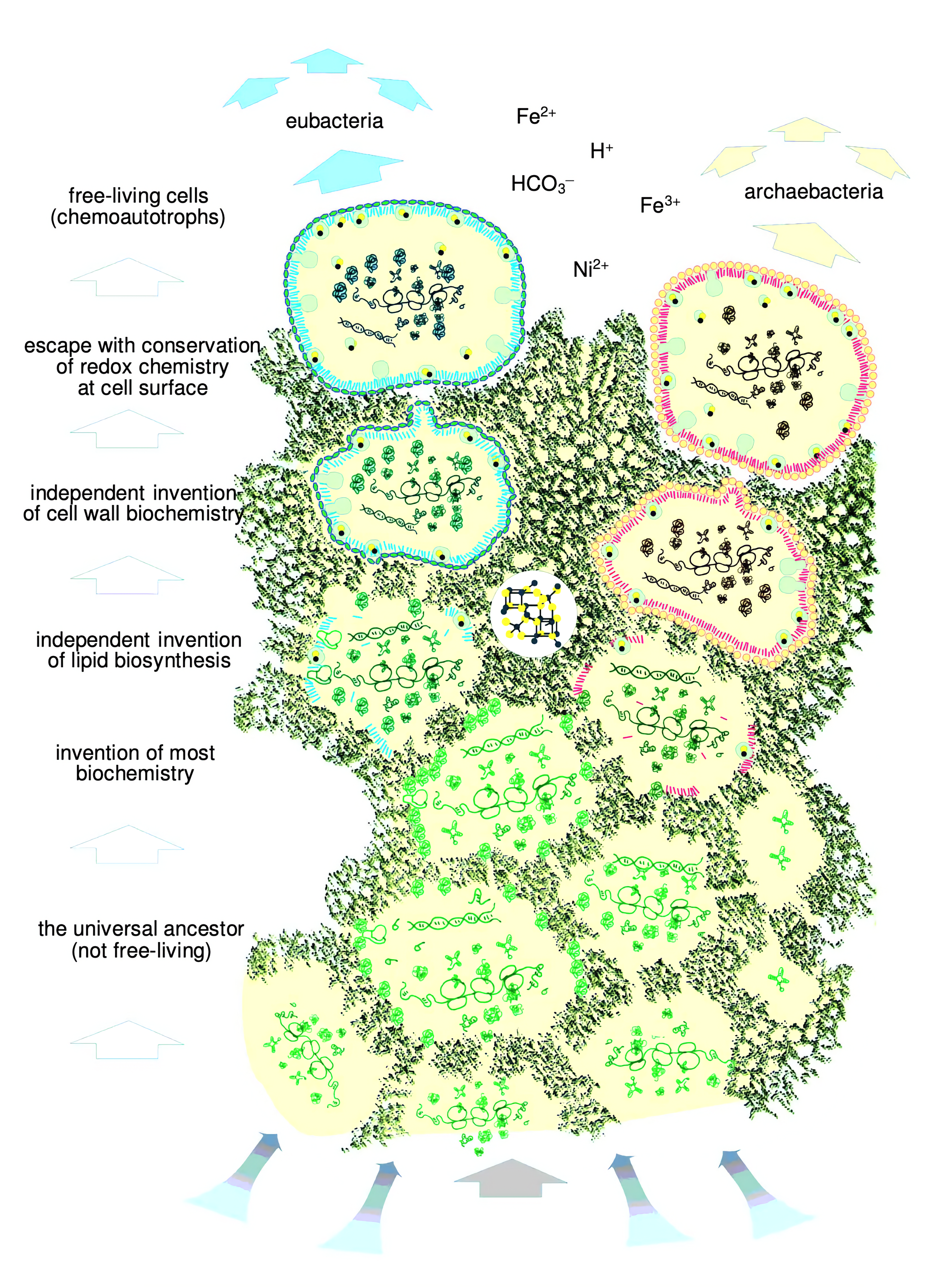
Obwohl diese Organismen bereits einen rudimentären Stoffwechsel und einen primitiven Genapparat besitzen, sind sie für viele ihrer Reaktionen immer noch stark von der katalytischen Aktivität der Eisen-Schwefel-Oberflächen abhängig. Im Laufe der Evolution entwickeln sie aber Enzyme welche die Eisen-Schwefel-Mineralien ersetzen, indem sie teils Eisen-Schwefel-Kerne in die Proteine mit einbauen. Aufgrund von Mikrofossilien und noch heute beobachtbarer physiologischen und genetischen Unterschieden zwischen Archaeen und Bakterien geht man davon aus, dass sich bevor Organismen eigene Zellmembranen entwickelt haben, Archaeen und Bakterien von einander abgespalten haben.
Ein Beispiel dafür ist, dass Archaeen und Bakterien stark unterschiedliche stark unterschiedliche Zellmembranen besitzen, was darauf zurückzuführen ist, dass sie jeweils unabhängig voneinander ihre eigenen Baupläne für Zellmembranen entwickelt haben.
Nun ausgestattet mit einer eigenen Zellmembran, welche die Molekül in sich konzentriert hält und vor äußeren Einwirkungen schützen kann, können diese Protozellen nun den Poren der Eisen-Schwefel-Schwämme entkommen und ihr Dasein als freilebende Organismen beginnen.
Entwicklung von Eukaryoten
Die Entwicklung von Eukaryoten, auch Eukaryogenese genannt, ist ein komplexes Thema, da Eukaryoten jeweils Merkmale von Archaeen und von Bakterien besitzen und gleichzeitig Merkmale, welche exklusiv in Eukaryoten zu sehen sind. Wo letzteres relativ einfach durch evolutionäre Neuerungen zu erklären ist, ist es schwer vorstellbar, dass ein Archaeon von alleine eine große Menge von bakteriellen Merkmalen entwickelt hat oder vice versa. Die Annahme, dass Eukaryoten von Prokaryoten abstammen, ist in sich selbst kontrovers, löst diese Annahme das Problem nicht, sondern verdoppelt es, somit müsste eine neue Lebensform gleichzeitig archaeische und bakterielle Merkmale entwickeln.
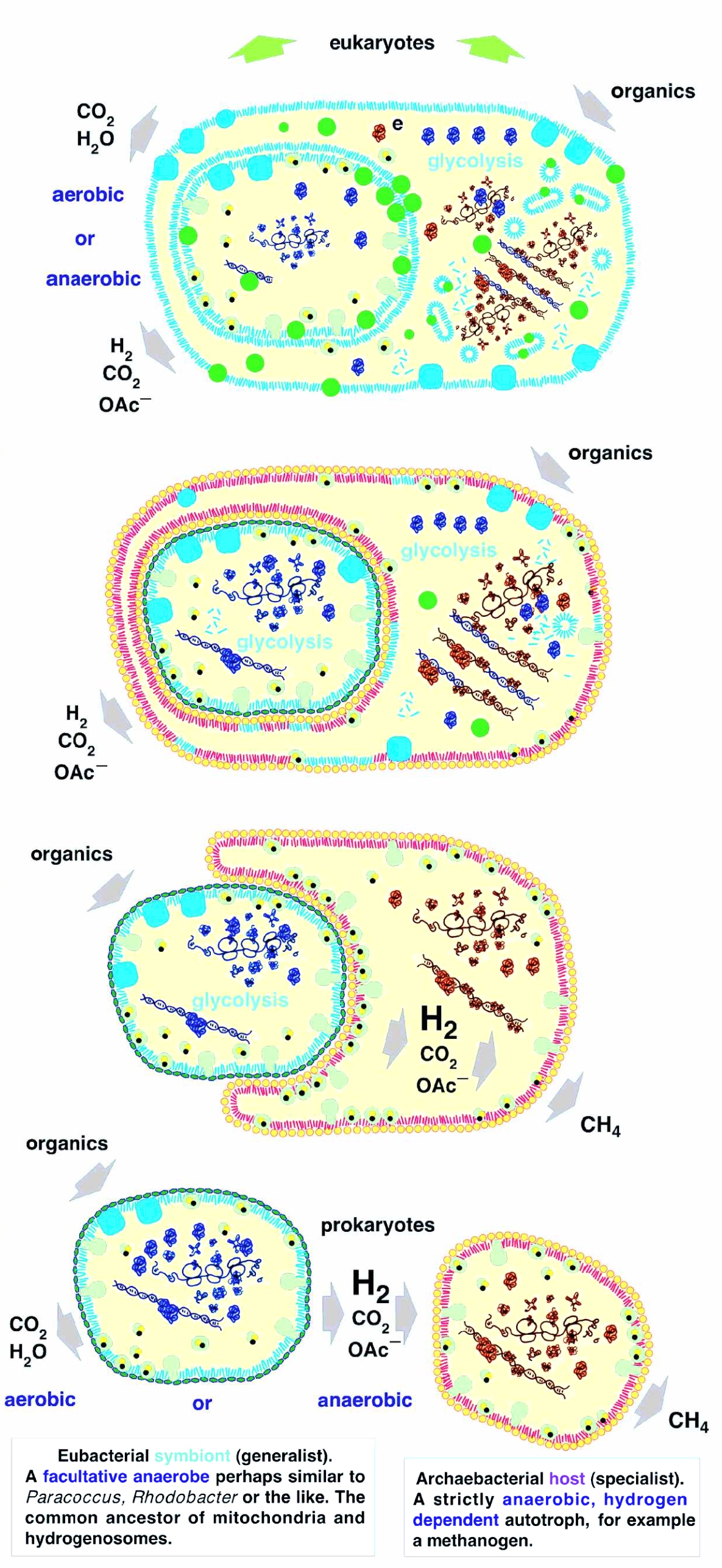
Das Modell, welches am besten zur derzeitigen Beweislage, bestehend aus Hunderten von Tausenden von Mikrofossilien und der Analyse von abertausenden sequenzierten Genomen von Archaeen, Bakterien und Archaeen, ist, dass ein Bakterium eine endosymbiotische Beziehung mit einem Archaeon eingegangen ist und später von diesem absorbiert wurde.
Der Symbiont ist ein Bakterium, welches anaerobe Gärung betreibt. Bei anaerober Gärung werden größere organische Moleküle aufgespalten und in ihre Bestandteile zerlegt, wobei elementarer Wasserstoff als ein Abfallprodukt anfällt.
Der Wirt hingegen ist ein chemoautotrophes Archaeon. In einem chemoautotrophen Stoffwechsel synthetisiert der Organismus komplexere Biomoleküle aus kleinen anorganischen Molekülen, wobei der Organismus seine Energie aus der chemischen Umsetzung von Stoffen selbst gewinnt. Für unser Szenario ist ebenfalls wichtig, dass der Wirt elementaren Wasserstoff als Elektronendonator und allgemein energiereichen Stoff für seinen Stoffwechsel benötigt.
Somit ist es logisch, dass das Archaeon und das Bakterium eine symbiotische Beziehung eingehen. Irgendwann nimmt das Archaeon das Bakterium in sich auf, um besser von dem von ihm produzierten Wasserstoff Gebrauch machen zu können, somit wird das letztere zu einem Endosymbionten. Jedoch benötigt das Bakterium immer noch organische Verbindungen von außen. Es besitzt jedoch keine Kohlenstoffimporteure, welche im Endosymbionten dafür verantwortlich sind, organische Verbindungen von außen in die Zelle zu schleusen. Also muss das Archaeon die Gene für diese Kohlenstoffimporteur-Proteine exprimieren. Es kommt zu einem horizontalen Gentransfer, einem Prozess, in welchem Gene aus einem Organismus in das Genom eines anderen Organismus übertragen werden. Dieser Transfer passiert hier, im Gegensatz zum vertikalen Gentransfer, welcher zwischen Eltern(-n) und Kind geschieht, zwischen zwei bereits bestehenden Organismen.
Neben diesen exprimiert der Wirt jedoch noch andere Gene des Endosymbionten, welche für Glykolyse und ATP-Produktion zuständig sind. Diese neuen Stoffwechselwege ersetzen auf lange Sicht die alten Stoffwechselwege des Archaeons.
Im letzten Schritt degeneriert das Bakterium von einem eigenen Organismus zu einer Endomembran, welche den Grundstein für die Entstehung eines Zellkerns legt. Währendessen werden weitere Gene aus dem Bakterium in das Archaeon transferiert, welche dazu führen, dass der Wirt nun eine bakterienähnliche Membran anstatt einer archaeischen Membran bildet. Ebenfalls bilden diese transferierten Gene die Grundlage für die Entstehung von Mitochondrien.